Sobrecruzamiento
Definición de sobrecruzamiento
El sobrecruzamiento es el intercambio de material genético entre cromátidas no hermanas de cromosomas homólogos durante la meiosis, lo que da como resultado nuevas combinaciones alélicas en las células hijas.
Cada célula diploide contiene dos copias de cada cromosoma, una derivada del gameto materno y la otra del gameto paterno. Estos pares de cromosomas, cada uno derivado de uno de los padres, se denominan cromosomas homólogos. Cuando los organismos diploides se reproducen sexualmente, primero producen gametos haploides a través de la meiosis. Durante la profase I de la meiosis, los cromosomas homólogos se alinean entre sí e intercambian material genético, de modo que algunos de los cromosomas resultantes son recombinantes, que contienen una mezcla de genes derivados de los cromosomas materno y paterno.
Cromosomas homólogos
La mayoría de los organismos que se reproducen sexualmente contienen dos tipos de células en su cuerpo: células haploides y diploides. Las células diploides se ven en la mayor parte del cuerpo y contienen dos copias de cada cromosoma. Por lo tanto, contienen dos genes para determinar la secuencia de casi todos los ARN o proteínas. Las células haploides suelen ser gametos masculinos o femeninos que se forman como resultado de la meiosis y se observan en los órganos sexuales. Contienen solo un juego completo de cromosomas y están diseñados para fusionarse con otro gameto haploide para producir un cigoto diploide, con un juego de cromosomas paterno y otro materno. Aunque codifican los mismos genes, sus secuencias de ADN pueden variar ligeramente entre sí. Por ejemplo, el cromosoma paterno puede contener la información genética para que el antígeno A esté presente en rojo.células sanguíneas, mientras que el cromosoma materno puede codificar el antígeno B. Estos dos antígenos son similares en función, pero difieren marginalmente entre sí en su secuencia de aminoácidos.
Todas las células de un organismo adulto se derivan del cigoto inicial y, por lo tanto, son genéticamente idénticas entre sí. Cuando este adulto se prepara para la reproducción, algunas células especializadas se someten a meiosis y producen cuatro células hijas haploides. Estas células hijas pueden madurar para formar gametos masculinos o femeninos.
Durante la meiosis, los cromosomas derivados del gameto materno y paterno pueden clasificarse independientemente unos de otros. En otras palabras, los cromosomas de la madre o del padre se pueden elegir al azar para cada gameto. Un espermatozoide humano puede contener los cromosomas 1, 3, 5, 7, 9, 21 y 23 de su progenitor femenino, y el resto puede surgir del progenitor masculino. De manera similar, un óvulo puede tener solo dos cromosomas del progenitor femenino y el resto del progenitor masculino.
Además, cada uno de estos cromosomas podría ser una mezcla recombinante de genes que surgen de los dos padres. Una de las cuatro células hijas haploides que surgen de la meiosis podría tener un cromosoma con el 80% de su secuencia del padre masculino y el 20% restante del padre femenino. En consecuencia, habrá otra celda hija con la relación inversa. Esto ocurre a través del cruzamiento, cuando los cromosomas homólogos replicados se colocan cerca unos de otros e intercambian segmentos entre sí.
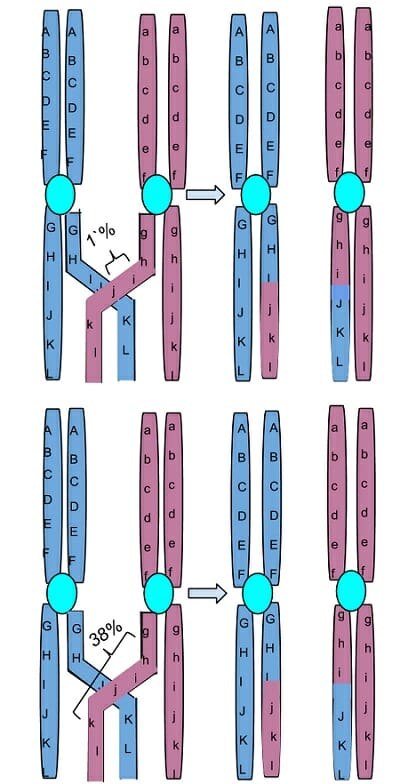
La imagen es una representación de un conjunto de cromosomas homólogos, con genes representados por diferentes letras del alfabeto. Los genes de un cromosoma se muestran con letras mayúsculas y los del par homólogo con letras minúsculas. Dos de las cuatro células hijas formadas después de este evento de cruzamiento tienen un cromosoma recombinante que no se deriva completamente de la madre ni del padre. La imagen también demuestra que es probable que los genes que se encuentran en estrecha proximidad física entre sí en el cromosoma se hereden juntos, mientras que los que están más alejados pueden clasificarse de forma independiente durante la meiosis.
Mapas de enlace
Los mapas de ligamiento son estimaciones de la distancia entre dos loci genéticos, basadas en la frecuencia de recombinación. Los loci de genes que están lejos unos de otros en el mismo cromosoma, o los que se encuentran en diferentes cromosomas, tienen un 50% de posibilidades de heredarse juntos y un 50% de probabilidades de ser clasificados de forma independiente. En función de la frecuencia con la que se heredan juntos, se pueden formar mapas de ligamiento, donde se muestra la posición de estos genes en relación con marcadores genéticos conocidos. Por ejemplo, si un marcador genético en el cromosoma X se hereda con frecuencia junto con un alelo particular de la calvicie de patrón masculino, se dice que forman parte del mismo grupo de ligamiento. Antes de la secuenciación de ADN de alto rendimiento, los mapas de ligamiento fueron las herramientas principales para comprender qué genes estaban presentes en un cromosoma en particular.
Profase I
La meiosis ocurre en dos etapas: meiosis I y II. La meiosis I, también conocida como división de reducción, es la serie de eventos que resulta en la formación de dos células hijas haploides. Al final de la división de reducción, el número de cromosomas se reduce a la mitad y cada una de las células hijas tiene solo un juego completo de cromosomas duplicados.
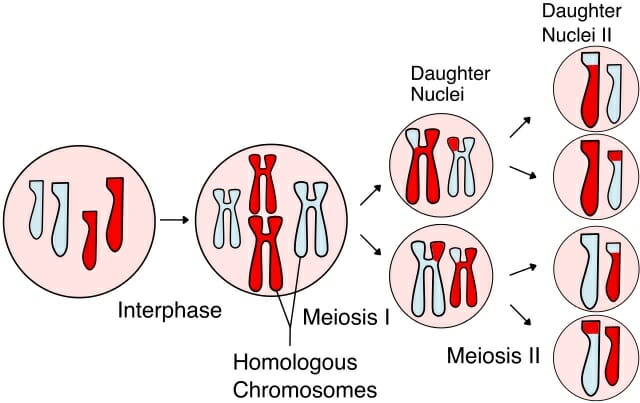
Durante la meiosis I, particularmente la profase I, ocurren varios eventos, lo que la convierte en una de las fases más largas de la meiosis. Marca el comienzo de la condensación cromosómica, donde los cromosomas duplicados con cromátidas hermanas adheridas se ven inicialmente como hilos largos y delgados. A medida que avanza la condensación, los cromosomas homólogos se unen debido a la similitud en la estructura y la posición del centrómero. Una estructura proteicaEl llamado complejo sinaptonemal también juega un papel importante. En este punto, los cromosomas están anclados a la envoltura nuclear. Ahora, la recombinación ocurre entre cromátidas no hermanas de cromosomas homólogos. Esto se observa microscópicamente como un evento de sobrecruzamiento entre cromosomas bivalentes (un par de dos cromosomas) con una estructura de tétrada (sus cromátidas hermanas duplicadas también son visibles).
Hacia el final de la profase I, los cromosomas homólogos ahora parecen «repelerse» entre sí. La envoltura nuclear ya no es claramente visible y la célula pasa a la metafase y la anafase para completar la primera etapa de la meiosis.
Mecanismo de sobrecruzamiento
A nivel molecular, el sobrecruzamiento comienza con una ruptura de doble hebra en una de las moléculas de ADN. Esta rotura de doble hebra puede ocurrir naturalmente a través de agentes como radiación o carcinógenos, o mediante la acción de proteínas específicas. Posteriormente, las exonucleasas, enzimas que eliminan nucleótidos del extremo 5 ‘del ADN, actúan sobre esta ruptura y eliminan tramos cortos de nucleótidos en la orientación 5’ -> 3 ‘de ambas cadenas. Esto conduce a dos regiones monocatenarias colgantes que se recubren con proteínas que catalizan la recombinación, también conocidas como recombinasas. Estas enzimas catalizan la invasión de regiones monocatenarias en secuencias que son adecuadas para el apareamiento de bases. La estrecha proximidad de las cromátidas no hermanas durante la profase I permite que esta región monocatenaria utilice la secuencia en el cromosoma homólogo.cromátida como plantilla. Esto conduce a que su hebra complementaria se desplace y se empareje de bases con la segunda región monocatenaria que fue inicialmente generada por la exonucleasa.
En última instancia, esto da como resultado el intercambio de dos hebras con la formación de una estructura en forma de cruz llamada unión de Holliday. Esto lleva el nombre del científico que propuso por primera vez que tal unión podría explicar tanto el sobrecruzamiento como otro fenómeno llamado conversión génica, donde un locus genético heterocigoto se vuelve homocigótico durante la división celular. Las uniones de Holliday también pueden verse microscópicamente como ‘quiasma’ hacia el final de la profase I, que continúan siendo visibles hasta el final de la anafase I. Las uniones de Holliday se estabilizan y resuelven a través de proteínas que modulan la manipulación genómica que se conocen como MSH4 y MSH5.
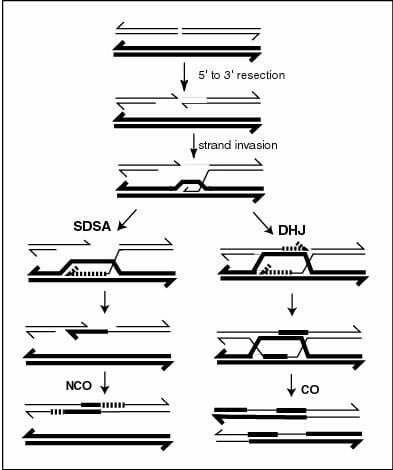
En la imagen, los eventos posteriores a la invasión de cadenas que conducen al sobrecruzamiento y la formación del sobrecruzamiento de Holliday se muestran a la derecha. Los eventos que no se cruzan, incluida la conversión génica, se muestran a la izquierda.
Funciones del sobrecruzamiento
Los organismos que se dividen solo asexualmente sin la posibilidad de tal recombinación sufren de una condición llamada Trinquete de Muller. Es decir, cada generación de esa especie contiene al menos tantas mutaciones genéticas como la generación anterior, si no más. En otras palabras, cuando toda la progenie es genéticamente idéntica entre sí, no hay margen para corregir errores genéticos o para que surjan combinaciones nuevas y beneficiosas.
El sobrecruzamiento aumenta la variabilidad de una población y previene la acumulación de combinaciones deletéreas de alelos, mientras que también permite que algunas combinaciones parentales se transmitan a la descendencia. De esta forma, existe un equilibrio entre mantener combinaciones alélicas potencialmente útiles y brindar la oportunidad de variación y cambio.
Términos de biología relacionados
- Citocinesis: división del contenido del citoplasma en dos células hijas al final de la mitosis o meiosis.
- Cariocinesis: división del contenido de un núcleo durante la división celular.
- Ploidía: número de conjuntos de cromosomas en una célula. Si bien los haploides (un grupo) y diploides (dos grupos) son los más comunes, algunos organismos pueden contener células triploides o incluso tetraploides.
- Cigoto: una célula diploide formada como resultado de la fusión de dos gametos haploides.